Propionil-CoA karboksilaza (PCC) je 750 kDa alfa (6) -beta (6)-dodekamer. (Samo približno 540 kDa je nativni enzim.[8]) Alfa podjedinice su raspoređene kao monomeri u centralnob beta-6 heksamernom jezgru. Spomenuto jezgro orijentirano je kao kratki cilindar s rupom duž svoje osi.
Alfa podjedinica PCC-a sadrži domene biotin-karboksilaza (BC) i biotinskog karboksilnog nosača proteina (BCCP). Domen poznat kao BT domen također se nalazi na alfa podjedinici i bitan je za interakcije sa beta podjedinicom. Lanac-8 antiparalelnog beta-bačvastog nabora ovog domena posebno je zanimljiv. Beta podjedinica ima aktivnost karboksiltransferaze (CT).[9]
a. Struktura himere RpPCCα-RdPCCβ, gledano niz osu trostruke simetrije. Domene u α i β podjedinicama u gornjoj polovini strukture imaju različite boje, a one u prvim α i β podjedinicama su označene. Podjedinice α i β u donjoj polovini obojene su u magenta i zeleno. Crvena strelica označava smjer gledanja ploče b. (b). Struktura himere RpPCCα-RdPCCβ, gledano niz osu dvostruke simetrije. Crveni pravougao označava područje detaljno prikazano na slici 2a. (c). Krio-EM rekonstrukcija HsPCC u rezoluciji 15 Å, gledano u istoj orijentaciji kao i ploča a. Atomski model himere uklopljen je u omotač krio-EM. (d). Krio-EM rekonstrukcija gledana u istoj orijentaciji kao panel b. Strelice označavaju promjenu položaja BCCP-a koja je potrebna da se uklopi u krio-EM mapu. Sve strukturne figure proizvedene su s PyMOL-om (www.pymol.org), a krio-EM slike Chimerom.[10] BC i CT mjesta udaljena su približno 55 Å, što ukazuje na to da se cijeli domen BCCP translocira tokom katalize karboksilacijapropionil-CoA.[10] Ovo daje jasne dokaze o ključnoj dimernosti interakcija između alfa i beta podjedinica.(a). Relativno pozicioniranje BC i CT aktivnih mjesta u holoenzimu. Prikazani su jedna α podjedinica i β2 dimer (β1 iz jednog sloja i β4 iz drugog sloja), a smjer gledanja je isti kao na slici 1b. Dva aktivna mjesta označena su zvijezdjcama, odvojenim udaljenošću od 55 Å. Prikazani su i vezani položaji ADP-a u kompleksu sa E. coli BC 18 i položaja CoA u kompleksu sa 12S podjedinicom transkarboksilaze 21. (b). Detalji interakcije između BCCP-biotina i C domena β podjedinice. Interakcije vezivanja vodika označene su isprekidanim crvenim linijama. N1′ atom biotina označen je kao 1′, vodikom vezan za karbonil glavnog lanca Phe397. (c). Molekulska površina CT aktivnog mjesta, prikazuje duboki usjek, gdje su vezane obje podloge. (d). Shema CT aktivne lokacije.[10]
Džep PCC-a za vezivanje biotina je hidrofoban i visoko konzerviran. Biotin i propionil-CoA vezuju se okomito jedan na drugi u oksijanionskom udubini, koja sadrži aktivno mjesto. Utvrđeno je da je omjer nativnog enzima i biotina jedan mol nativni enzim na četiri mola biotina. Smatra se da je N1 biotina baza aktivnog mjesta.
Mutageneza usmjerena na lokus na D422 pokazuje promjenu specifičnosti supstrata za mjesto vezivanja propionil-CoA, što ukazuje na značaj ovog ostatka u katalitskoj aktivnosti PCC-a.[11] U 1979., inhibicijom fenilglioksala utvrđeno je da fosfatna grupa iz propionil-CoA ili ATP reagira esencijalnim ostatkom arginina na aktivnom mjestu tokom katalize.[12] Kasnije (2004.) predloženo je da arginin-338 služi za orijentaciju karboksifosfatnog intermedijara za optimalnu karboksilaciju biotina.[13]
KM utvrdila je da su vrijednosti ATP, propionil-CoA i bikarbonata 0,08 mM, odnosno 0,29 mM i 3,0 mM. Izoelektrična tačka pada na pH 5,5. Strukturni integritet PCC -a očuvan je u temperaturnom rasponu od –50 do 37 stepeni Celzijusa i rasponu pH od 6,2 do 8,8. Pokazalo se da je optimalni pH između 7,2 i 8,8 bez vezanog biotina, Sa biotinom, optimalni pH je 8,0-8,5.[14]
Enzimski mehanizam
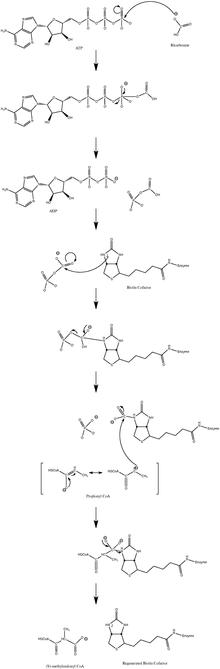
Uobičajeni mehanizam katalitske reakcije uključuje karbanionski međuproizvod i ne prolazi kroz usklađen proces.[15] Slijedeća slika prikazuje vjerovatni put.
Vjerovatni mehanizam PCC
Pokazalo se da je reakcija blago reverzibilna, pri niskom protoku propionil-CoA.[16]
Izozimi
Ljudi eksprimiraju sljedeće dvije propionil-CoA karboksilaznih izoenzima:
Aktivnost PCC je najosjetljiviji pokazatelj statusa biotina koji je do sada testiran. U budućim studijama trudnoće upotreba podataka o aktivnosti PCC-a limfocita trebala bi se pokazati vrijednom za procjenu statusa biotina.[20]
Unutargenska komplementacija
Kada više kopija polipeptida kodiranog genom tvori agregat, ova proteinska struktura naziva se multimer. Kada se multimer formira od polipeptida koje proizvode dva različita mutantnaalela određenog gena, mješoviti multimer može pokazati veću funkcionalnu aktivnost od nepomiješanih multimera koje tvori svaki od mutanata. U tom slučaju, fenomen se naziva intragenska komplementacija.
PCC je heteropolimer, sastavljen od α i β podjedinica u strukturi α6β6. Mutacija u PCC -u, bilo u α (PCCα) ili β podjedinici (PCCβ) može uzrokovati propionsku acidemiju kod ljudi. Kada su različite ćelijske linije kožnih fibroblasta s kožnomdefektnom PCCβ spojene u parovima, β heteromultimerni protein, nastao kao njena posljedica često je pokazivao viši nivo aktivnosti nego što bi se očekivalo na osnovu aktivnosti roditeljskih enzima.[21] Ovaj nalaz intragenske komplementacije) pokazao je da multimerna struktura PCC-a omogućava kooperativne interakcije između sastavnih monomera PCCβ koji mogu generirati funkcionalniji oblik holoenzima.
Regulacija
Propionil-CoA karboksilazom
a. Karbamazepin (antiepileptik): značajno snižava nivo enzima u jetri[22]
^Kalousek F, Darigo MD, Rosenberg LE (1980). "Isolation and characterization of propionyl-CoA carboxylase from normal human liver. Evidence for a protomeric tetramer of nonidentical subunits". The Journal of Biological Chemistry. 255 (1): 60–65. PMID6765947.
^Diacovich L, Mitchell DL, Pham H, Gago G, Melgar MM, Khosla C, Gramajo H, Tsai SC (2004). "Crystal Structure of theβ-Subunit of Acyl-CoA Carboxylase: Structure-Based Engineering of Substrate Specificity†,‡". Biochemistry. 43 (44): 14027–14036. doi:10.1021/bi049065v. PMID15518551.
^Wolf B, Kalousek F, Rosenberg LE (1979). "Essential arginine residues in the active sites of propionyl CoA carboxylase and beta-methylcrotonyl CoA carboxylase". Enzyme. 24 (5): 302–306. doi:10.1159/000458679. PMID510274.
^Ugarte M, Pérez-Cerdá C, Rodríguez-Pombo P, Desviat LR, Pérez B, Richard E, Muro S, Campeau E, Ohura T, Gravel RA (1999). "Overview of mutations in thePCCA and PCCB genes causing propionic acidemia". Human Mutation. 14 (4): 275–282. doi:10.1002/(SICI)1098-1004(199910)14:4<275::AID-HUMU1>3.0.CO;2-N. PMID10502773.
^Desviat LR, Pérez B, Pérez-Cerdá C, Rodríguez-Pombo P, Clavero S, Ugarte M (2004). "Propionic acidemia: Mutation update and functional and structural effects of the variant alleles". Molecular Genetics and Metabolism. 83 (1–2): 28–37. doi:10.1016/j.ymgme.2004.08.001. PMID15464417.
^Deodato, F.; Boenzi, S.; Santorelli, F. M.; Dionisi-Vici, C. (2006). "Methylmalonic and propionic aciduria". American Journal of Medical Genetics Part C. 142C (2): 104–112. doi:10.1002/ajmg.c.30090. PMID16602092. S2CID21114631.
^Rodríguez-Pombo P, Pérez-Cerdá C, Pérez B, Desviat LR, Sánchez-Pulido L, Ugarte M. Towards a model to explain the intragenic complementation in the heteromultimeric protein propionyl-CoA carboxylase. Biochim Biophys Acta. 2005;1740(3):489-498. doi:10.1016/j.bbadis.2004.10.009
^McKeon C, Wolf B (1982). "Magnesium and magnesium adenosine triphosphate activation of human propionyl CoA carboxylase and beta-methylcrotonyl CoA carboxylase". Enzyme. 28 (1): 76–81. doi:10.1159/000459088. PMID6981505.
^Zhang H, Boghigian BA, Pfeifer BA (2010). "Investigating the role of native propionyl-CoA and methylmalonyl-CoA metabolism on heterologous polyketide production inEscherichia coli". Biotechnology and Bioengineering. 105 (3): 567–573. doi:10.1002/bit.22560. PMID19806677. S2CID659042.